
Eco-epidemiological studies to develop integrated vector surveillance of leishmaniasis vectors in the Americas

 ,
, Abstract
We developed an integrated vector surveillance (IVS) proposal for cutaneous leishmaniasis (CL) and visceral leishmaniasis (VL) in the Americas, based on eco-epidemiological studies conducted by researchers of the Leishmaniasis Research Network of Argentina. For CL, the transmission was explained in the framework of the edge effect, the increase of vectors and risk of exposure at ecotones and environmental interfaces, and typified as ephemeral, transient, or permanent edges, supporting a cost-effective IVS strategy for early warning of CL outbreaks through an environmental modification alert network, which includes multiple sources of information and actors. In relation to VL, the earliest colonization sites and spatial distribution were explained by modeling and forecasting the most likely hotspots, persistent in time and space, and modulated by environmental variables. Therefore, for VL, a scalar strategy of critical site selection is proposed from a “city” scale based on secondary sources such as remote sensing for the definition of possible areas to monitor and intervene, a scale of restriction from possible to most likely areas through local knowledge, and a “focal site” scale of trap placement through field observation; in this way, IVS activities are carried out at a few sites of the urban landscape and allow a sustainable program.
Keywords
INTRODUCTION
In the Pan American Health Organization/World Health Organization manuals, integrated vector management (IVM) is characterized as an information-based decision-making procedure that optimizes the cost-effectiveness and cost-benefit of vector control. This definition refers to a logical process for the efficient use of resources, with a sustainable strategy, of low socio-environmental impact which involves one or more synergistic interventions for one or more vectors[1].
On the other hand, leishmaniasis in the Americas needs the development, evaluation, validation, and implementation of new IVM strategies, due to the magnitude of its impact on public health, its status as an emerging or re-emerging tropical neglected disease, and the relatively low residuality of the chemical interventions currently used for its control[2]. Cutaneous leishmaniasis (CL) generates focal epidemic outbreaks that can affect hundreds of people; in its cutaneous-mucosal manifestation; it is mutilating and involves the risk of mortality due to secondary upper respiratory tract infections or suicide associated with facial ulcers and difficulty in speech and socialization. Visceral leishmaniasis (VL), a disease of urban transmission in rapid geographic dispersion, presents very high lethality without specific chemotherapy, and the intervention of asymptomatic domestic dogs as a reservoir introduced new problems for its control[3-6]. The incidence records of VL and CL in the Americas by country are periodically updated in the leishmaniasis reports of the Pan American Health Organization.
In relation to the appropriate conceptual framework for approaching and structuring the development of IVM, epidemiology, ecology, and panoramic epidemiology, based on the analysis of collective health events in time and space, attempt to provide the where and when of transmission. Meanwhile, this same knowledge, through a multi-scale and multi-dimensional approach, integrating physical, biological, social, cultural, economic, and political variables from eco-epidemiology, attempts to describe the why. This last comprehensive theoretical approach, as a contribution to instrumental and operational aspects, is close to the objective of integrated management. However, for many vector-borne diseases, more IVM proposals have been generated in relation to control interventions than to surveillance strategies. Therefore, this article develops a proposal for integrated vector surveillance for CL and another for VL in the Americas, based on some of the work and collaborations carried out by the Leishmaniasis Research Network in Argentina and the experience of 30 years of fieldwork on the subject.
CUTANEOUS LEISHMANIASIS
In relation to CL in a continental analysis, at a macro-scale, but with data disaggregated by a municipality, different clusters of environmental and socio-economic variables were observed, associated with different levels of incidence of human CL and epidemic events. The clusters with the higher casuistry and the larger spatial extension, such as the Amazonian cluster, are related to environments that have been little modified, and activities of intrusion into the forest or deforestation with the migration of susceptible people. However, in all clusters, a spatial relationship is observed between environmental modification events and epidemic events[5], which justifies that, as a preventive strategy, environmental management and surveillance could be a sustainable and effective strategy[7].
In Argentina, of the 46 species of Phlebotominae recorded thus far, four are the most important for CL transmission, being proven or incriminated vectors of Leishmania braziliensis, the main circulating parasite, although there are increasing records of L. amazonensis and sporadic reports of L. panamensis and L. guyanensis[8,9]. According to the ecoregion, the dominant populations at the forest-rural-urban interfaces are the Phlebotominae Nyssomyia neivai in the western and eastern subtropical area, Ny. whitmani in the extreme northwest, and Migonemyia migonei and the Evandromyia complex in the central region[10,11].
At the mesoscale, in the hyper-endemic area of the northwest, the largest recent epidemic outbreaks of CL are recorded on both sides of the border of Argentina and Bolivia in the same years during 1998, 2002, and 2015-2018, possibly as a consequence of shared climatic and agro-industrial deforestation events at the same time on both sides of the border[12,13]. This “perfect storm” conjunction forms the so-called eco-epidemiological momentum[14], and in this case, its association with environmental modification activity is reflected in the higher incidence in males of active working age[15]. Therefore, for northwestern Argentina, based on the presence and abundance of competent vectors in areas of human-vector interaction, a stratified risk map was developed, where epidemic outbreak risk sites were also related to areas of environmental interface and intense anthropic modification of land use. The predictive distribution map for Ny. neivai and Mg. migonei, as mentioned, the two main vector species in the area, showed association with “precipitation in warmest quarter” and “mean temperature in coldest quarter”[16]. Both climatic variables may be related to the inter-annual difference in the area of tree cover in the ecotone, thus modifying the total breeding area for Phlebotominae[17], which oviposit in shady, humid, and rich in organic matter sites but not flooded or with intense traffic[18].
At the microscale transmission site scale, the results of the other scales are explained by the edge effect, which generates a focal increase in vector abundance at environmental interfaces created by massive deforestation or even by small “islands” produced by forest clearing, even with no settlement of domestic animals or human on the intervened side[19,20]. However, when this new blood supply exists, as with the installation of poultry houses, the arrival of vectors from the extra-domicile to the peri-domicile is significantly increased, and new breeding sites for sandflies are generated close to the animal farms[18]. In the conceptual framework of the edge effect, many CL epidemic events can also be explained: (a) by the generation of very short-lived edges due to intrusion or ephemeral camps in areas of primary or secondary vegetation[20-23] by progressive peri-urban deforestation with the presence of contiguous domestic spaces and land occupation[24-26]; and (b) by urbanization with stable peripheral edges, intra-urban green spaces, or riparian forests[27-31]. Indirectly, outbreaks can also be explained by the movement of susceptible people to a non-residential area for forest-associated work[32]; concentration of vectors, reservoirs, and people due to a flood, fire, or other catastrophic events[33,34]; translocation of vectors due to the disappearance of their food sources[35]; or synanthropic wild reservoirs that may increase the possibility of vector contact at edges[36,37].
The edges, in turn, may prove to be an area of periodic colonization. Nyssomyia neivai in the hyperendemic region shows at microscale a cross-correlation of abundance between the extra-domestic and peri-domestic areas every two adult-to-adult cycles. This correlation would indicate the possibility of sink populations in the anthropized habitats, periodically colonized by source populations of parental flying adults from the more conserved area. This metapopulation dynamics would also be consistent with the possibility of climatic modulation by increasing the suitable breeding area of Phlebotominae referred above, since the abundance of Ny. neivai is positively associated with temperature and relative humidity with cyclical time lags up to one year prior to capture, which would correspond to windows of favorable conditions for the previous cohorts. In relation to human incidence and vector abundance, the correlation shows a shift between the peak of the probable time of infection of human cases (winter) and the peak of Ny. neivai females most likely to be infected (late autumn), due to the intrinsic incubation time of Leishmania in humans[11,12,17].
From an environmental epidemiological point of view, these previous arguments allowed the categorization of four cycles: (a) wild with the transmission in primary or residual vegetation; (b) wild with eventual peri-domestic transmission by disturbance of residual vegetation patches; (c) wild with peri-domestic transmission by contiguity with residual vegetation; and (d) peri-domestic in rural environments, ruralized peri-urban, or urban-rural interface[38]. When the time of exposure and permanence of humans in relation to the ecotone and the risk activities that generate this exposure are added to this analytical discrimination based on the edge effect, epidemic outbreaks can be classified into three main epidemiological patterns
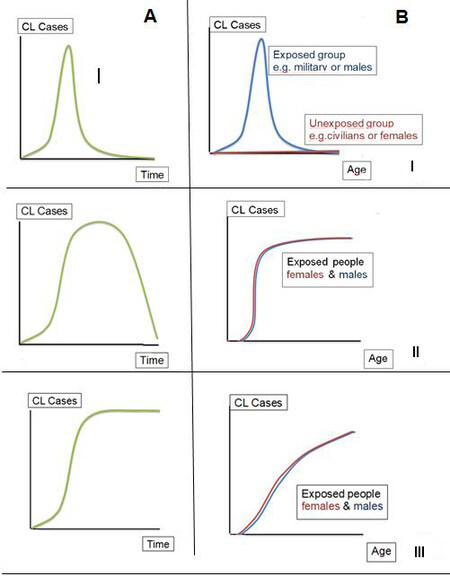
Figure 1. Patterns of human cutaneous leishmaniasis incidence over time (A) and CL incidence by age and gender (B) according to the most common scenarios of: (I) ephemeral edge; (II) transient edge; and (III) permanent edge.
(I) Ephemeral edge: This is due to the intrusion, transit, or brief stay in the forest, with an incidence curve over time in the shape of a narrow bell, and there is a common point source in time and space. Incidence curves by sex and age group are also bell-shaped, as only the age group, sex, and profession exposed are infected due to military practices, prospecting and research, or legal and illegal extractive activities.
(II) Transient edge: This is due to the mobile deforestation front, temporary camp, or natural disaster, and the incidence curve over time is a longer bell-shaped curve than in Pattern (I) according to the time exposure has been sustained. There are sigmoid-similar infection incidence curves by age group for both sexes, with early exponential growth and a stationary phase due to saturation of susceptible people. The exposure disappears during progressive deforestation, land occupation, and housing development of new suburbs when the edge increases its distance from the domestic area or such area turns to unsuitable conditions for vector colonization with the absence of uncovered land surface.
(III) Permanent edge: This develops in front of ecotones and interfaces with stable edges and sigmoidal or sinusoidal incidence curves over time with periodic bells at sites of strongly seasonal transmission. The curves by sex increase similarly with age due to cumulative risk over time, but they could turn stationary in the older age groups (survival stages).
In any case, infection curves identified by sex or age may differ when the disease is recorded due to differences in the probability of clinical expression. On the ephemeral edge, cultural practices can alter these patterns, for example, in extractive activities with the participation of the whole family as an active or accompanying party, as occurs in the harvesting of wild fruits or migration through illegal passes. In the transient and permanent borders, there may be age displacements due to the protection of younger age groups with mosquito nets or intra-domiciliary rest during twilight and night hours or displacements due to domestic activities with restricted exposure for a defined age group and sex during hours of vector activity, such as water collection, firewood collection, domestic animal management, etc.[14].
The described works, typifications, and the referential framework of the edge effect support a proposal for a cost-effective integrated surveillance strategy for the early warning of CL epidemic outbreaks in the Americas through an environmental modification alert network, which includes multiple sources of information and actors. To this aim, with existing information, the surveillance coordinators should stratify the risk areas according to the vulnerability of the exposed population and current or potential edges generated by anthropogenic activities or climatic events. This stratification would be dynamic, with periodic inputs from surveillance data and the results of focal investigations.
Once the areas have been determined, as well as the intensity with which surveillance should be carried out at each site according to the probability of epidemic events, the map of actors would be identified for each spatial scale. This map integrates the degree of legal responsibility of each actor, their agency capacity to modify or mitigate the risk, or otherwise the need for their empowerment, which in turn implies describing the flow, frequency, and mode of exchange of information in both directions. In this way, the network should integrate, among others: (A) actors at the international level, such as development project funding agencies, which should include CL risk assessment and monitoring among the requirements and evaluation of impact; (B) actors at the national level or first subnational jurisdictions, in addition to the health sector, areas involved in environmental change or population interaction with edges such as forest regulation agencies, meteorological and remote sensing monitoring agencies for natural disasters, armed forces that carry out activities in the forest, environmental non-governmental organizations; (C) actors at the municipal level such as those responsible for urban development, civil defense, labor unions, and primary healthcare agents; and (D) focal actors, individual volunteers, media, and social networks.
This network involves identifying sites of risk of CL outbreaks with epidemiological, entomological, and environmental data to stratify these risks and actors at each scale and record by local knowledge the most exposed people by specific practices, their schedule, seasonality, or socio-environmental and occupational vulnerability[39-43]. Thus, before a change in land use, a legal or illegal activity, or an environmental alteration in a project or in progress that may involve an edge, this information should be immediately reported and evaluated in situ with entomological surveys; (re)training and sensitization of primary healthcare agents in the early identification of suspected cases and the referral and follow-up of healthcare system agents for the proper diagnosis, management, and notification of cases and social protection agencies agents to mitigate the impact on the quality of family or group life. If the event is occurring, a focused study and the provision of inputs for case management should be carried out; if it may occur, baseline studies and follow-up should be carried out. In all cases, it is expected that prevention or mitigation measures can be proposed by shifting risk to places or times of lower vector abundance or the generation of physical (deforested sanitary belts) and/or chemical barriers[44,45].
VISCERAL LEISHMANIASIS
Visceral leishmaniasis in Argentina and its vectors have records prior to 2000[46]; however, since the spread of urban transmission from Brazil[4], Lutzomyia longipalpis was recorded for the first time in Argentina in 2004 in urban environments[47], and the first human case was reported in 2006[48], both in the northwest on the border with Paraguay. Dispersal continued westward, reaching 22°30′ S 63°50′ W in 2013[49,50] and southward[43,51-54], reaching 31°18' S 58°00' W in 2017[55], such that in both directions, the urbanized vector traveled about 800-850 km. The cases of canine VL and human VL by vector-borne transmission are currently spreading but distributed spatially in a discontinuous way[56], so the surveillance system might be able to detect the early stages of colonization of Lu. longipalpis, the main vector of L. infantum in urban settings, anticipating the occurrence of cases, first in dogs and later in humans, to raise awareness in both the veterinary and human health systems, the community, and the vector control agencies according to the local spatial and temporal patterns of the vector[3,57,58]. The incidence tends to stabilize after some time in localities in the southernmost latitudes of vector transmission distribution, but which continue to generate cases at a lower frequency, maintain canine prevalence, or may destabilize and generate a new outbreak[59].
The urban VL surveillance, in order to be a sustainable strategy, given the geographical extension to be covered, should be focused on the “within the city” riskier sites instead of blanket activities that try to cover all the space of the city with the same intensity, and in the same sense, it should be periodic at the time of highest risk, not continuous. Furthermore, besides the known distribution and surveillance of
However, Lu. longipalpis, still the main urban reservoir in most the Americas, has been shown to be a species where different populations have different biological characteristics including pheromones, dispersal ability, urban adaptation, and potentially different vectorial capacity[61]. In this sense, in Argentina, the different populations of Lu. longipalpis registered high rates of genetic diversity, grouping into three haplogroups, two exclusive to Argentina and the third shared with Brazil. The spatial distribution of these haplogroups suggests that the colonization event in Argentina’s territory occurred through different events at different sites and in different locations, giving rise to hybrids between the new “urban invaders” and the previous “rural residents”. This double adaptive contribution may thus generate new adaptive capacities or vectorial capacity[50,62,63]. In this regard, related to less urbanized populations of vectors, it should be noted that, although currently predominantly urban with the dog as a reservoir[64], L. infantum infection has been found in the non-human primate Alouatta caraya, with vectors distributed at the base and top of trees used by primates as roosts[65,66].
However, from the point of the need for surveillance of a selection strategy of sites at locality scale of highest probability of initial vector ocurrence, it was observed from the first description of urban infestation in Argentina that the persistence of sites with high vector abundance over time through a highly discontinuous matrix of infestation[11,26,67,68]. This persistence in time and space allowed us to model the distribution of intraurban Lu. longipalpis, confirming the presence of hotspots of vector abundance in the cities of Posadas, Corrientes, Santo Tomé, Clorinda, and Puerto Iguazu with a spatial autocorrelation of 400-600 m[69-76]. These patterns could correspond to metapopulation dynamics with the hotspots acting as source populations; thus, during the most appropriate months for Phlebotominae, the vectors could disperse to other sites of the city by extending the risk area for a few months. Therefore, targeted intervention in these persistent “islands” of Lu. longipalpis abundance could have a cost-effective impact on the distribution and risk of vector transmission throughout the locality, not only as a surveillance strategy but also as an intervention strategy The possibility of characterizing the environmental variables associated with these sites, based on models developed by different analytical approaches, allows the integration of knowledge of vector ecology to propose a strategy for the selection of the critical sites[77,78]. The lower is the number of sites selected, most likely it is to be colonized by Lu. longipalpis (monitoring) or to function as source populations of the vector (preventive control), and the higher is the cost-effectiveness of the integrated vector surveillance strategy. This proposal, as for cutaneous leishmaniasis, should also be implemented in a successive scalar manner from macro- to microscale [Figure 2]. At the “city” spatial level, secondary sources such as remote sensing for environmental indicators, census data for demographic indicators, and human or veterinary case surveillance system data, if available, can be used to determine likely hotspots. With these probable areas visualized as patches or islands on a map or satellite image, local actors, according to their knowledge of the socio-environmental geography of their city, would be able to define at the macrohabitat level within the possible critical areas previously indicated the probable critical areas of more restricted dimensions.
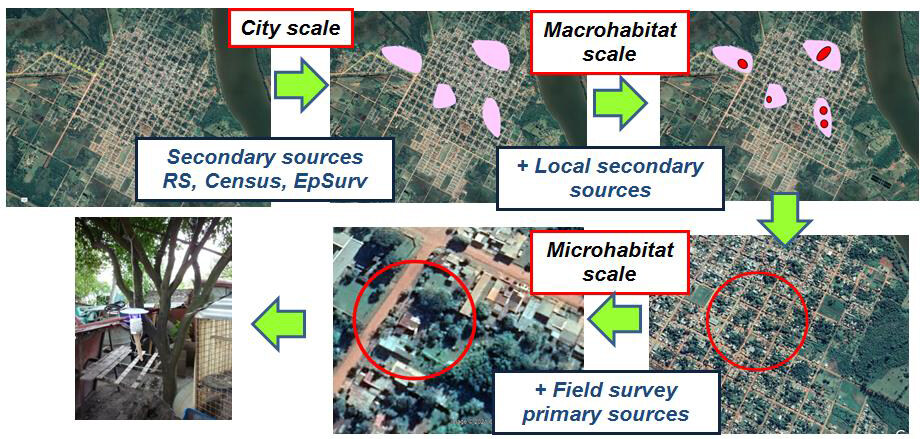
Figure 2. Scalar surveillance strategy for urban visceral leishmaniasis, sources of information and selection of possible critical sites at city scale (pink patches), probable critical sites at macrohabitat scale (red spot), and critical sites for installation of surveillance at microhabitat scale (red circle). Source of images: Google Earth and own photographs.
Finally, with the probable critical areas defined, the microhabitats can be identified, thus selecting the critical sites - peri-domicile and domicile - where to place the vector traps, using primary data derived from the field survey and primary healthcare information.
The selection criteria for each scale would be defined in operational manuals that can be adapted to regional characteristics. A user-friendly interface of the algorithm for the generation of probable critical areas from satellite images should be developed for its management by technicians at the municipal level. The manual should also define the minimum size of locality where this strategy can be applied, since in the very small ones, a “census” monitoring could imply the same effort where a hotspot scalar selection or selection of sites by local knowledge would be enough.
CONCLUSION
The analysis from the theoretical framework of eco-epidemiology, the integrated vector management information-based decision-making procedure tools, and the One Health intersectoral coordination operation, applied to leishmaniasis research in Argentina, generated sustainable proposals for integrated vector surveillance for leishmaniases in the Americas. For CL, a multiscale environmental surveillance network strategy was developed, while a scalar critical site selection system was developed for urban VL. In both cases, different levels of agents should be involved with different degrees of commitment and responsibility in prevention and focal intervention, promoting the empowerment of local communities, in addition to knowing the local seasonal dynamics of the vectors in order to perform or intensify surveillance during periods of higher abundance. Both strategies are in the stage of field validation and evaluation of their co-effectiveness.
DECLARATIONS
AcknowledgmentsTo all the members of the Leishmaniasis Research Network of Argentina (REDILA), and collaborators at federal, provincial and local jurisdictions.
Authors’ contributionsContributed to the conceptualization and writing of the article: Salomón OD, Quintana MG.
Availability of data and materialsData and materials were previously published, references are cited in the article.
Financial support and sponsorshipNo specific funding was received for the writing of this article, the sources of funding for the research reviewed are cited in the respective referenced articles.
Conflicts of interestAll authors declared that there are no conflicts of interest.
Ethical approval and consent to participateNot applicable.
Consent for publicationNot applicable.
Copyright© The Author(s) 2022.
REFERENCES
1. OPS. Documento operativo de aplicación del manejo integrado de vectores adaptado al contexto de las Américas. Available from: https://iris.paho.org/bitstream/handle/10665.2/51760/9789275320990_spa.pdf [Last accessed on 18 May 2022].
2. OPS. Manual de procedimientos para vigilancia y control de las leishmaniasis en las Américas. Available from: https://iris.paho.org/handle/10665.2/50524 [Last accessed on 18 May 2022].
3. Salomón OD, Mastrángelo AV, Santini MS, Ruvinsky S. Visceral leishmaniasis: paths that converge and divide. Available from: https://www.researchgate.net/publication/281771111_Visceral_leishmaniasis_Paths_that_converge_and_divide [Last accessed on 18 May 2022].
4. Salomón OD, Feliciangeli MD, Quintana MG, Afonso MM, Rangel EF. Lutzomyia longipalpis urbanisation and control. Mem Inst Oswaldo Cruz 2015;110:831-46.
5. Maia-Elkhoury ANS, Magalhães Lima D, Salomón OD, et al. Interaction between environmental and socioeconomic determinants for cutaneous leishmaniasis risk in Latin America. Rev Panam Salud Publica 2021;45:e83.
6. OPS. Métodos de vigilancia entomológica y control de los principales vectores en las Américas. Available from: https://iris.paho.org/handle/10665.2/55241 [Last accessed on 18 May 2022].
7. Orellano PW, Vazquez N, Salomon OD. Cost-effectiveness of prevention strategies for American tegumentary leishmaniasis in Argentina. Cad Saude Publica 2013;29:2459-72.
8. Moya SL, Giuliani MG, Santini MS, Quintana MG, Salomón OD, Liotta DJ. Leishmania infantum DNA detected in phlebotomine species from Puerto Iguazú City, Misiones province, Argentina. Acta Trop 2017;172:122-4.
9. Moya SL, Szelag EA, Manteca-Acosta M, Quintana MG, Salomón OD. Update of the phlebotominae fauna with new records for Argentina and observations on leishmaniasis transmission scenarios at a regional scale. Neotrop Entomol 2022;51:311-23.
10. Salomón OD, Rosa JR, Stein M, et al. Phlebotominae (Diptera: Psycodidae) fauna in the Chaco region and cutaneous leishmaniasis transmission patterns in Argentina. Mem Inst Oswaldo Cruz 2008;103:578-84.
11. Quintana MG, Fernández MS, Salomón OD. Distribution and abundance of phlebotominae, vectors of leishmaniasis, in Argentina: spatial and temporal analysis at different scales. J Trop Med 2012;2012:652803.
12. Salomón OD, Quintana MG, Rosa JR. Ecoepidemiología de la leishmaniasis cutánea en Argentina. Available from: https://www.researchgate.net/profile/Oscar-Salomon/publication/285844101_Eco-epidemiology_of_cutaneous_leishmaniasis_in_Argentina/links/57f78e1b08ae886b89834363/Eco-epidemiology-of-cutaneous-leishmaniasis-in-Argentina.pdf [Last accessed on 18 May 2022].
13. Mollinedo JS, Mollinedo Z, Magne M, Gironda WJ, Salomón ÓD. Leishmaniasis en Bolivia. Comprehensive review and current status in Tarija, in the border with Argentina. Biomedica 2020;40:45-61.
14. Salomon OD. Instructions on how to make an outbreak of American cutaneous leishmaniasis. J Trop Med Health 2019;3:146.
15. Sosa-Estani S, Segura EL, Gomez A, et al. [Cutaneous leishmaniasis in Northern Argentina: identification of risk factors in a case-cohort study of three municipalities in Salta]. Rev Soc Bras Med Trop 2001;34:511-7.
16. Quintana M, Salomón O, Guerra R, De Grosso ML, Fuenzalida A. Phlebotominae of epidemiological importance in cutaneous leishmaniasis in northwestern Argentina: risk maps and ecological niche models. Med Vet Entomol 2013;27:39-48.
17. Salomón OD, Wilson ML, Munstermann LE, Travi BL. Spatial and temporal patterns of phlebotomine sand flies (Diptera: Psychodidae) in a cutaneous leishmaniasis focus in northern Argentina. J Med Entomol 2004;41:33-9.
18. Manteca-Acosta M, Cavia R, Utgés ME, Salomón OD, Santini MS. Peridomestic natural breeding sites of Nyssomyia whitmani (Antunes and Coutinho) in an endemic area of tegumentary leishmaniasis in northeastern Argentina. PLoS Negl Trop Dis 2021;15:e0009676.
19. Salomon OD, Rossi GC, Spinelli GR. Ecological aspects of phebotomine (Diptera, Psychodidae) in an endemic area of tegumentary leishmaniasis in the northeastern Argentina, 1993-1998. Mem Inst Oswaldo Cruz 2002;97:163-8.
20. Quintana MG, Salomón OD, De Grosso MS. Distribution of phlebotomine sand flies (Diptera: Psychodidae) in a primary forest-crop interface, Salta, Argentina. J Med Entomol 2010;47:1003-10.
21. Salomón OD, Sosa Estani S, Canini L, Córdoba Lanús E. Leishmaniosis tegumentaria en un área con niveles epidémicos de transmisión, Salta, Argentina, 1998. Medicina (B Aires) 2001;61:284-90.
22. Salomón OD, Acardi SA, Liotta DJ, et al. Epidemiological aspects of cutaneous leishmaniasis in the Iguazú falls area of Argentina. Acta Trop 2009;109:5-11.
23. Szelag EA, Rosa JR, Quintana MG, Salomon OD. Temporal distribution of, and effect of anthropic modifications on, phlebotomine populations in the Chaco Bioregion, Argentina. Med Vet Entomol 2018;32:206-15.
24. Salomón OD, Sosa Estani S, Monzani AS, Studer C. Brote epidémico de leishmaniosis tegumentaria en Puerto Esperanza, provincia de Misiones, 1998. Medicina (Buenos Aires) 2001;61:385-90.
25. Salomón OD, Sosa-Estani S, Ramos K, et al. Tegumentary leishmaniasis outbreak in Bella Vista City, Corrientes, Argentina during 2003. Mem Inst Oswaldo Cruz 2006;101:767-74.
26. Santini MS, Fernández MS, Cavia R, Salomón OD. Co-occurrence and seasonal and environmental distributions of the sandflies Lutzomyia longipalpis and Nyssomyia whitmani in the city of Puerto Iguazú, northeastern Argentina. Med Vet Entomol 2018;32:197-205.
27. Salomon OD, Bogado De Pascual M, Molinari ML, Verri V. Study of a cutaneous leishmaniasis outbreak in General Vedia, Province of Chaco, 1996. Rev Inst Med Trop Sao Paulo 2001;43:99-104.
28. Salomón OD, Quintana MG, Zaidenberg M. Urban distribution of phlebotominae in a cutaneous leishmaniasis focus, Argentina. Mem Inst Oswaldo Cruz 2008;103:282-7.
29. Salomon OD. Addressing the recent dispersion of urban visceral leishmaniasis in the border of Argentina, Brazil, Paraguay+ Uruguay+ Bolivia - Project IDRC. Rev Inst Adolfo Lutz 2018;77:e1756.
30. Thomaz-Soccol V, Gonçalves AL, Piechnik CA, et al. Hidden danger: unexpected scenario in the vector-parasite dynamics of leishmaniases in the Brazil side of triple border (Argentina, Brazil and Paraguay). PLoS Negl Trop Dis 2018;12:e0006336.
31. Fernández MS, Manteca-Acosta M, Cueto GR, Cavia R, Salomón OD. Variation of the phlebotominae (Diptera: Psychodidae: Phlebotominae) assemblage in response to land use changes in an endemic area of leishmania transmission in northeast Argentina. J Med Entomol 2020;57:1735-47.
32. Salomon OD, Zaidenberg M, Burgos R, Heredia VI, Caropresi SL. American cutaneous leishmaniasis outbreak, Tartagal city, province of Salta, Argentina, 1993. Rev Inst Med Trop Sao Paulo 2001;43:105-8.
33. Salomón OD, Sosa Estani S, Drí L, et al. Leishmaniosis tegumentaria en Las Lomitas, provincia de Formosa, Argentina, 1992-2001. Medicina (B Aires) 2002;62:562-8.
34. Salomón OD, Orellano PW, Lamfri M, et al. Phlebotominae spatial distribution asssociated with a focus of tegumentary leishmaniasis in Las Lomitas, Formosa, Argentina, 2002. Mem Inst Oswaldo Cruz 2006;101:295-9.
35. Salomón OD, Quintana MG, Flores I, et al. Phlebotominae sand flies associated with a tegumentary leishmaniasis outbreak, Tucumán Province, Argentina. Rev Soc Bras Med Trop 2006;39:341-6.
36. Acardi SA, Rago MV, Liotta DJ, Fernández-Duque E, Salomon OD. Leishmania Viannia DNA detection by PCR-RFLP and sequencing in free-ranging owl monkeys (Aotus azarai azarai) from Formosa, Argentina. Vet Parasitol 2013;193:256-9.
37. Fernández MS, Fraschina J, Acardi S, et al. Assessment of the role of small mammals in the transmission cycle of tegumentary leishmaniasis and first report of natural infection with Leishmania braziliensis in two sigmodontines in northeastern Argentina. Parasitol Res 2018;117:405-12.
38. Salomón OD, Orellano PW, Quintana MG, et al. Transmisión de la leishmaniasis tegumentaria en Argentina. Medicina (B Aires) 2006;66:211-9.
39. Mastrángelo AV, Salomón OD. Trabajo forestal y Leishmaniasis Cutánea: Un análisis social centrado en el riesgo para el N de Misiones (Argentina). Available from: https://www.researchgate.net/publication/277790388_Trabajo_forestal_y_Leishmaniasis_Cutanea_Un_analisis_social_centrado_en_el_riesgo_para_el_N_de_Misiones_Argentina [Last accessed on 18 May 2022].
40. Mastrángelo AV, Salomón OD. Contribución de la antropología a la comprensión ecoepidemiológica de la Leishmaniasis Tegumentaria Americana en las “2000 hectáreas”, Puerto Iguazú, Misiones, Argentina. Available from: https://rasp.msal.gov.ar/index.php/rasp/article/view/423 [Last accessed on 18 May 2022].
41. Fuenzalida AD, Quintana MG, Salomón OD, de Grosso MS. Hourly activity of Lutzomyia neivai in the endemic zone of cutaneous leishmaniasis in Tucumán, Argentina: preliminary results. Mem Inst Oswaldo Cruz 2011;106:635-8.
42. Fernández MS, Lestani EA, Cavia R, Salomón OD. Phlebotominae fauna in a recent deforested area with American tegumentary leishmaniasis transmission (Puerto Iguazú, Misiones, Argentina): seasonal distribution in domestic and peridomestic environments. Acta Trop 2012;122:16-23.
43. Salomón OD, Mastrángelo AV, Santini MS, Liotta DJ, Yadón ZE. La eco-epidemiología retrospectiva como herramienta aplicada a la vigilancia de la leishmaniasis en Misiones, Argentina, 1920-2014. Rev Panam Salud Publica 2016;40:29-39.
44. Bates PA, Depaquit J, Galati EA, et al. Recent advances in phlebotomine sand fly research related to leishmaniasis control. Parasit Vectors 2015;8:131.
45. Acosta M, Santini MS, Pérez AA, Salomón OD. Evaluation of efficacy of impregnated curtains in experimental hen houses as a phlebotomine control tool in northeast Argentina. Med Vet Entomol 2017;31:161-6.
46. Salomón OD, Rossi G, Sosa Estani S, Spinelli G. Presencia de Lutzomyia longipalpis y situación de la leishmaniosis visceral en Argentina. Medicina (B Aires) 2001;61:174-8.
47. Salomón OD, Orellano PW. Lutzomyia longipalpis in Clorinda, Formosa province, an area of potential visceral leishmaniasis transmission in Argentina. Mem Inst Oswaldo Cruz 2005;100:475-6.
48. Salomon O, Sinagra A, Nevot M, et al. First visceral leishmaniasis focus in Argentina. Mem Inst Oswaldo Cruz 2008;103:109-11.
49. Salomón OD, Rosa JR, Fabiani M, San Miguel SR, Szelag EA, et al. Distribución de Lutzomyia longipalpis en el chaco argentina, 2010. Medicina (B Aires) 2011;71:225-30.
50. Quintana MG, Pech-May A, Fuenzalida AD, et al. Lutzomyia longipalpis (Diptera: Psychodidae) Argentina-Bolivia border: new report and genetic diversity. Mem Inst Oswaldo Cruz 2019;114:e190184.
51. Salomón OD, Ramos LK, Quintana MG, Acardi SA, Santini MS, et al. Distribución de vectores de leishmaniasis visceral en la provincia de Corrientes. Medicina (B Aires) 2009;69:625-30.
52. Salomón OD, Fernández MS, Saavedra S, Montiel M, Ramos MA, et al. Distribución de Lutzomyia longipalpis en Mesopotamia Argentina, 2010. Medicina (B Aires) 2011;71:22-6.
53. Salomón OD, Basmajdian Y, Fernández MS, Santini MS. Lutzomyia longipalpis in Uruguay, first report and potential of visceral leishmaniasis transmission. Mem Inst Oswaldo Cruz 2011;106:381-2.
54. Salomón OD, Basmajdian Y, Fernández MS, Santini MS. Lutzomyia longipalpis in Uruguay: the first report and the potential of visceral leishmaniasis transmission. Mem Inst Oswaldo Cruz 2011;106:381-2.
55. Szelag EA, Parras MA, Fabiani M, Rosa JR, Salomón OD. Incipient colonisation of lutzomyia longipalpis in the city of resistencia, province of Chaco, Argentina (2010-2012). Mem Inst Oswaldo Cruz 2014;109:488-91.
56. Santini MS, Acosta MM, Utgés ME, Aldaz ME, Salomón OD. Presence of lutzomyia longipalpis and nyssomyia whitmani in Entre Rios, Argentina. Rev Inst Med Trop Sao Paulo 2018;60:e55.
57. Gould TI, Perner MS, Santini MS, Saavedra SB, Bezzi G, et al. Leishmaniasis visceral en la Argentina: notificación y situación vectorial (2006-2012). Medicina (B Aires) 2013;73:104-10.
58. López K, Tartaglino LC, Steinhorst II, Santini MS, Salomon OD. Risk factors, representations and practices associated with emerging urban human visceral leishmaniasis in Posadas, Argentina. Biomedica 2016;36:51-63.
59. Santini MS, Salomón OD, Acardi SA, Sandoval EA, Tartaglino L. Lutzomyia longipalpis behavior and control at an urban visceral leishmaniasis focus in Argentina. Rev Inst Med Trop Sao Paulo 2010;52:187-91.
60. Lamattina D, Berrozpe PE, Casas N, et al. Twice upon a time: the progression of canine visceral leishmaniasis in an Argentinean city. PLoS One 2019;14:e0219395.
61. Salomón OD, Quintana MG, Bezzi G, Morán ML, Betbeder E, Valdéz DV. Lutzomyia migonei as putative vector of visceral leishmaniasis in La Banda, Argentina. Acta Trop 2010;113:84-7.
62. Salomón OD, Araki AS, Hamilton JG, Acardi SA, Peixoto AA. Sex pheromone and period gene characterization of lutzomyia longipalpis sensu lato (Lutz & Neiva) (Diptera: Psychodidae) from Posadas, Argentina. Mem Inst Oswaldo Cruz 2010;105:928-30.
63. Pech-May A, Ramsey JM, González Ittig RE, et al. Genetic diversity, phylogeography and molecular clock of the lutzomyia longipalpis complex (Diptera: Psychodidae). PLoS Negl Trop Dis 2018;12:e0006614.
64. Moo-Llanes DA, Pech-May A, de Oca-Aguilar ACM, Salomón OD, Ramsey JM. Niche divergence and paleo-distributions of lutzomyia longipalpis mitochondrial haplogroups (Diptera: Psychodidae). Acta Trop 2020;211:105607.
65. Acardi SA, Liotta DJ, Santini MS, Romagosa CM, Salomón OD. Detection of leishmania infantum in naturally infected lutzomyia longipalpis (Diptera: Psychodidae: Phlebotominae) and canis familiaris in Misiones, Argentina: the first report of a PCR-RFLP and sequencing-based confirmation assay. Mem Inst Oswaldo Cruz 2010;105:796-9.
66. Martínez MF, Santini MS, Kowalewski MM, Salomón OD. Phlebotominae in peri-domestic and forest environments inhabited by alouatta caraya in northeastern Argentina. Med Vet Entomol 2019;33:367-74.
67. Martínez MF, Kowalewski MM, Giuliani MG, Acardi SA, Salomón OD. Molecular identification of Leishmania in free-ranging black and gold howler monkeys (alouatta caraya) in northeastern Argentina. Acta Trop 2020;210:105534.
68. Salomón OD, Quintana MG, Bruno MR, Quiriconi RV, Cabral V. Visceral leishmaniasis in border areas: clustered distribution of phlebotomine sand flies in Clorinda, Argentina. Mem Inst Oswaldo Cruz 2009;104:801-4.
69. Bravo AG, Quintana MG, Abril M, Salomón OD. The first record of lutzomyia longipalpis in the Argentine northwest. Mem Inst Oswaldo Cruz 2013;108:1071-3.
70. Fernández MS, Salomón OD, Cavia R, Perez AA, Acardi SA, Guccione JD. Lutzomyia longipalpis spatial distribution and association with environmental variables in an urban focus of visceral leishmaniasis, Misiones, Argentina. Acta Trop 2010;114:81-7.
71. Fernández MS, Santini MS, Cavia R, et al. Spatial and temporal changes in lutzomyia longipalpis abundance, a leishmania infantum vector in an urban area in northeastern Argentina. Mem Inst Oswaldo Cruz 2013;108:817-24.
72. Santini MS, Fernández MS, Pérez AA, Sandoval AE, Salomón OD. Lutzomyia longipalpis abundance in the city of Posadas, northeastern Argentina: variations at different spatial scales. Mem Inst Oswaldo Cruz 2012;107:767-71.
73. Santini MS, Utgés ME, Berrozpe P, et al. Lutzomyia longipalpis presence and abundance distribution at different micro-spatial scales in an urban scenario. PLoS Negl Trop Dis 2015;9:e0003951.
74. Gómez-Bravo A, German A, Abril M, Scavuzzo M, Salomón OD. Spatial population dynamics and temporal analysis of the distribution of lutzomyia longipalpis (Lutz & Neiva, 1912) (Diptera: psychodidae: phlebotominae) in the city of Clorinda, Formosa, Argentina. Parasit Vectors 2017;10:352.
75. Berrozpe P, Lamattina D, Santini MS, Araujo AV, Utgés ME, Salomón OD. Environmental suitability for lutzomyia longipalpis in a subtropical city with a recently established visceral leishmaniasis transmission cycle, Argentina. Mem Inst Oswaldo Cruz 2017;112:674-80.
76. Berrozpe PE, Lamattina D, Santini MS, Araujo AV, Torrusio SE, Salomón OD. Spatiotemporal dynamics of lutzomyia longipalpis and macro-habitat characterization using satellite images in a leishmaniasis-endemic city in Argentina. Med Vet Entomol 2019;33:89-98.
77. Quintana MG, Santini MS, Cavia R, et al. Multiscale environmental determinants of leishmania vectors in the urban-rural context. Parasit Vectors 2020;13:502.
Cite This Article
Export citation file: BibTeX | RIS
OAE Style
Salomón OD, Quintana MG. Eco-epidemiological studies to develop integrated vector surveillance of leishmaniasis vectors in the Americas. One Health Implement Res 2022;2:45-55. http://dx.doi.org/10.20517/ohir.2022.09
AMA Style
Salomón OD, Quintana MG. Eco-epidemiological studies to develop integrated vector surveillance of leishmaniasis vectors in the Americas. One Health & Implementation Research. 2022; 2(2): 45-55. http://dx.doi.org/10.20517/ohir.2022.09
Chicago/Turabian Style
Salomón, Oscar Daniel, María Gabriela Quintana. 2022. "Eco-epidemiological studies to develop integrated vector surveillance of leishmaniasis vectors in the Americas" One Health & Implementation Research. 2, no.2: 45-55. http://dx.doi.org/10.20517/ohir.2022.09
ACS Style
Salomón, OD.; Quintana MG. Eco-epidemiological studies to develop integrated vector surveillance of leishmaniasis vectors in the Americas. One. Health Implement. Res. 2022, 2, 45-55. http://dx.doi.org/10.20517/ohir.2022.09
About This Article
Copyright

Data & Comments
Data


 Cite This Article 5 clicks
Cite This Article 5 clicks

 Like This Article 5
likes
Like This Article 5
likes






Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.